


基底神经节传入
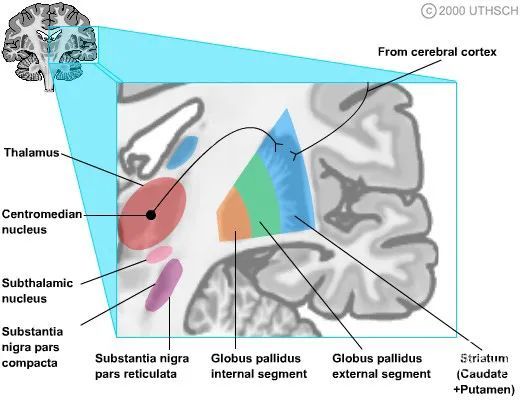
基底神经节传入。为简单起见,在本图中和后续图中,尾状核和壳核仅由壳核表示,因为这两个区域具有相似的连接。
纹状体是基底神经节传入的主要接收者。这些兴奋性传入来自整个大脑皮层和丘脑的椎板内核(主要是中央中核和束旁核)。
来自不同皮质区域的投射是分开的,因此额叶主要向尾状头和壳核投射;顶叶和枕叶投射到尾状核体;和颞叶投射到尾状核尾。
初级运动皮层和初级躯体感觉皮层主要投射到壳核,而前运动皮层和辅助运动区投射到尾状核头。其他皮质区域主要投射到尾状核。
因此,沿着尾状核的 C 形范围,尾状细胞从附近的皮层区域接收输入信号。扩大的尾状核头部放大了从额叶皮质到尾状核的投射信号。此外,伏隔核(腹侧纹状体)接收来自边缘皮质的大量传入信号。
在基底神经节的运动区,有一个类似于初级运动皮层的运动小脑。因此,来自中央旁小叶前部(M1 的一部分,包含代表腿部和躯干的区域)内侧壁的纤维投射支配着纹状体靠近中央前回背侧表面的受体区域。(M1 的一部分,其中包含代表手臂和手的区域)。类似地,来自中央前回(M1 中包含代表面部的区域)侧面的投射纤维支配着代表手臂和手部区域旁的部分。这种投射的地形保持在基底神经节的固有通路中。
基底神经节传出
基底神经节的主要输出结构是苍白球内部段 (GPint) 和黑质网状部 (SNr) 。这两种结构都产生 GABAergic(γ-氨基丁酸能),对目标产生抑制作用。GPint 通过两个纤维束投射到多个丘脑结构:豆状襻和豆核束。
处理来自运动皮层和躯体感觉皮层的感觉运动信息的环路投射到腹前核 (VA) 和腹外侧 (VL) 核。
处理其他新皮质信息的循环投射到背内侧核 (DM)、板内核和腹前核 (VA) 的部分。
SNr 投影到上丘,这涉及眼球运动,以及 VA/VL 丘脑核。
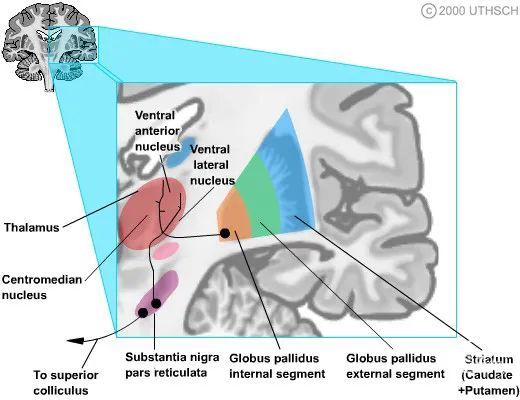
基底神经节传出
基底神经节内在连接
基底神经节的内部存在许多内部链接,相互联通。
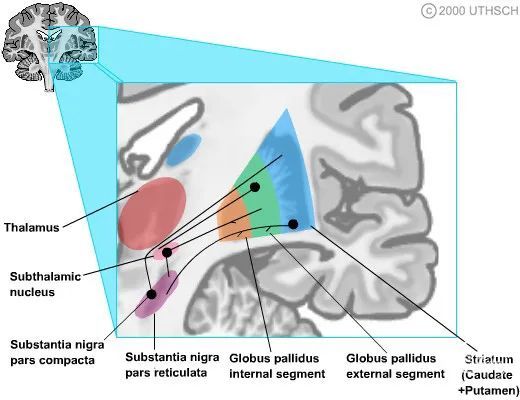
基底神经节内在连接
1、纹状体通路是纹状体和苍白球两个节段之间的 GABA 能抑制性连接。
2、纹状体黑质通路是纹状体和黑质网状部 (SNr)之间的 GABA 能抑制性连接。
3、苍白球外部节段与丘脑底核形成 GABA 能抑制性连接。
4、底丘脑核产生谷氨酸能,与苍白球和黑质网状部 (SNr)产生兴奋性连接。该通路是基底神经节内在通路中唯一的纯兴奋性通路。
5、黑质纹状体通路在纹状体神经元上形成多巴胺能突触。正如我们将在下面看到的,这是一个混合的途径,在一些纹状体神经元上有兴奋效应,在另一些上有抑制效应。
基底神经节中的两条通路处理信号
有两种不同的途径通过基底神经节处理信号:直接途径和间接途径。这两种途径对丘脑目标结构具有相反的净效应。直接通路的兴奋具有兴奋丘脑神经元的净效应(进而与皮层神经元产生兴奋性连接)。间接途径的激发具有抑制丘脑神经元的净效应(使它们无法激发运动皮层神经元)。基底神经节的正常功能显然涉及这两种途径的活动之间的适当平衡。一种假设是直接通路选择性地促进大脑皮层中适应当前任务的某些运动(或认知)程序,而间接通路同时抑制竞争性运动程序的执行。直接和间接途径之间的平衡被破坏会导致锥体外系综合征的运动功能障碍(见下文)。
直接途径。尽管直接和间接通路的连接模式相对简单,但系统中抑制连接的优势可以使对功能电路的理解变得复杂且不直观。
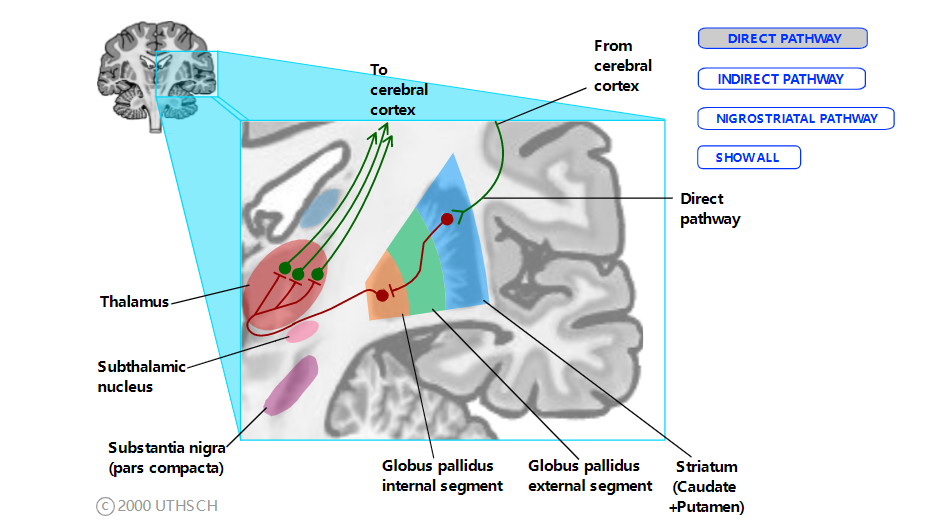
直接途径。实线代表直接途径。(GPi 的输出对两种途径都是通用的。)绿线代表兴奋性连接,红线代表抑制性连接。单击单个路径名称以单独查看每个路径。
直接途径始于纹状体中的细胞,这些细胞与 GPint 中的细胞建立抑制性连接。GPint 细胞反过来对丘脑中的细胞产生抑制性连接。因此,GPint 神经元的放电抑制了丘脑,使丘脑不太可能激发新皮质。然而,当直接通路纹状体神经元激活时,它们会抑制 GPint 神经元的活动。这种抑制解除了丘脑神经元的抑制(即解除丘脑神经元的抑制),使它们能够激发皮层。因此,由于纹状体和 GPint 以及 GPint 和丘脑之间通路的“双负”,刺激直接通路纹状体神经元的最终结果是兴奋运动皮层。
将其视为乘法方程,其中兴奋连接 (E) 等于 +1,抑制连接 (I) 等于 –1:

因为这两个负数相互抵消。
间接途径。间接途径始于纹状体中的一组不同的细胞。
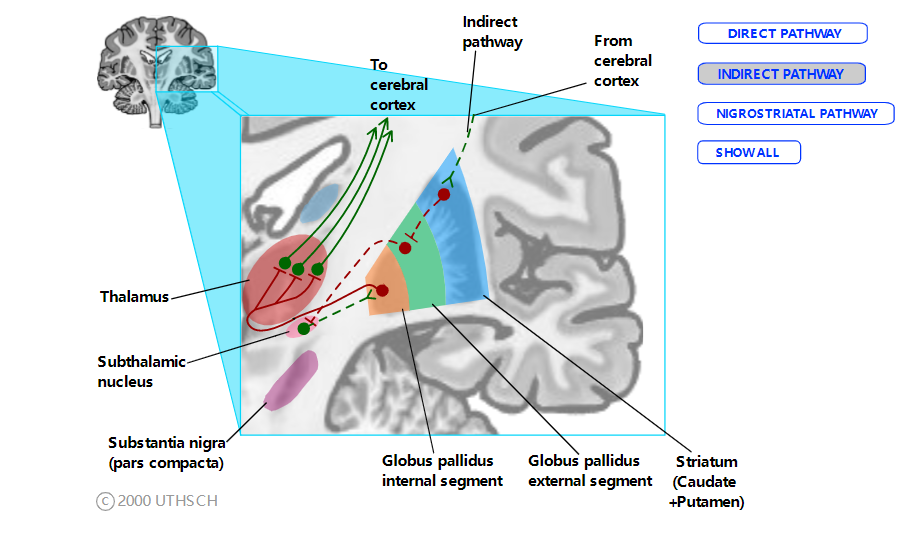
间接途径,虚线代表间接途径。(GPi 的输出对两种途径都是通用的。)绿线代表兴奋性连接,红线代表抑制性连接。单击单个路径名称以单独查看每个路径。
这些神经元与苍白球外段 (GPext) 建立抑制性连接。GPext 神经元与丘脑底核中的细胞建立抑制性连接,进而与 GPint 中的细胞建立兴奋性连接。(请记住,底丘脑-GPint 通路是内在基底神经节回路中唯一的纯兴奋性通路。)正如我们之前看到的,GPint 神经元对丘脑神经元进行抑制性连接。要查看间接途径激活的净效应,让我们从 GPint 向后工作。当 GPint 细胞活跃时,它们会抑制丘脑神经元,从而使皮层不那么活跃。当丘脑底神经元放电时,它们会增加 GPint 神经元的放电率,从而增加对皮层的净抑制。GPext 神经元的放电会抑制丘脑下神经元,从而使 GPint 神经元不那么活跃并解除对丘脑的抑制。然而,当间接通路纹状体神经元活跃时,它们会抑制 GPext 神经元,从而解除对丘脑下神经元的抑制。随着丘脑底神经元自由放电,GPint 神经元抑制丘脑,从而对运动皮层产生净抑制。
再想想乘法的类比:
![]()
因为等式中有 3 个负数,所以净效应为负。
因此,由于兴奋、抑制和去抑制的复杂序列,皮层刺激直接 通路的净效应是进一步兴奋皮层(正反馈回路),而皮层刺激间接通路的净效应是抑制皮层(负反馈回路)。据推测,基底神经节的功能与这两种途径之间的适当平衡有关。运动皮层神经元必须激发适当的直接通路神经元以进一步增加其自身的放电,并且它们必须激发适当的间接通路神经元,这些神经元将抑制其他不适应手头任务的运动皮层神经元(见下文)。
黑质纹状体投射
调节直接和间接途径的一个重要途径是多巴胺能——黑质纹状体投射从黑质致密部到纹状体。直接通路纹状体神经元具有 D1 多巴胺受体,可响应多巴胺使细胞去极化。相比之下,间接通路纹状体神经元具有 D2 多巴胺受体,可响应多巴胺使细胞超极化。因此黑质纹状体通路具有激发直接通路同时抑制间接通路的双重作用。由于这种双重效应,黑质纹状体通路的激发具有通过两种途径激发皮层的净效应,即通过激发直接通路(其本身对皮层具有净兴奋作用)和抑制间接通路(从而解除对净抑制作用的抑制)皮层上的间接途径)。
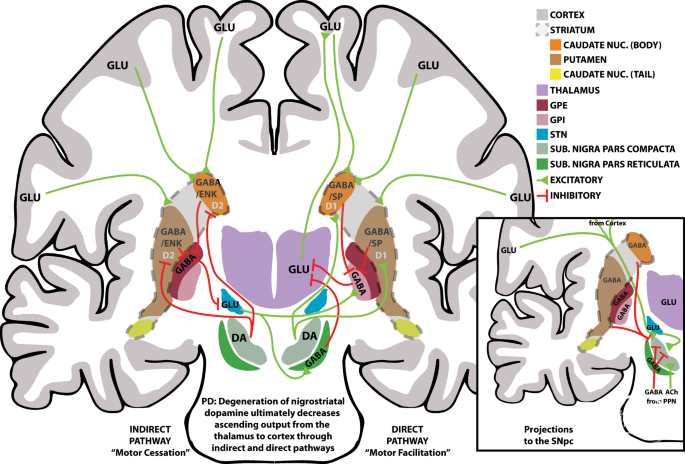
A schematic overview of the primary motor circuits in the basal ganglia, the indirect (left) and direct (right) pathways. Note, pathways crossing sides does not imply decussation, rather the contralateral connections separate the indirect and direct pathways. Excitatory connections are depicted in green with triangle ends, inhibitory connections are depicted in red with “T” ends. Not all connections are depicted, including but not limited to, all connections from thalamus to cortex, all connections from cortex to striatum, connections to/from caudate nucleus (tail), connections from cortex to brainstem, and inputs to SNpc (pictured in the inset). Inset: a schematic overview of inputs to SNpc found in literature. The PPN is located caudal to the substantia nigra and inputs are depicted as such. Inputs from the caudate nucleus (tail) is not pictured, and not all inputs from the cortex are depicted. Merging of signals from the cortex or caudate/putamen are done for illustrative purposes. D1; D1 receptors; D2; D2 receptors; DA dopamine; ENK enkephalin; GLU glutamate; NUC nucleus; PPN pedunculopontine nucleus; SP substance P; SUB substantia.
基底节的功能
运动功能
基底神经节在运动控制中的功能尚不清楚。似乎基底神经节通过调节存储在运动皮层和运动层次结构中的其他地方的运动程序,参与了练习运动行为的启用和自主运动的启动。因此,自主运动不是在基底神经节中开始的(它们是在皮层中开始的);然而,基底神经节的正常功能似乎是必要的,以便运动皮层将适当的运动命令传递到层次结构的较低级别。
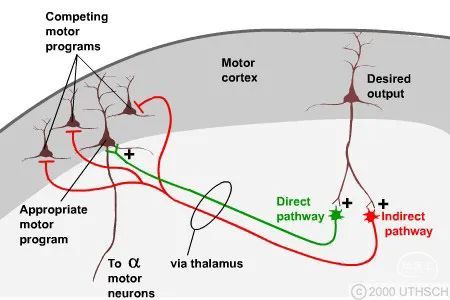
基底神经节和运动皮层形成一个处理循环,由此基底神经节通过直接通路启用存储在运动皮层回路中的适当运动程序,并通过间接通路抑制竞争性运动程序。根据从皮层传递的所需运动输出选择合适的运动程序。请注意,为清楚起见,直接和间接通路的复杂回路被示意性地绘制为单个神经元。
回想一下,基底神经节的主要输出是从 GPint(或 SNr)到丘脑(或上丘)的抑制性连接。对猴子眼球运动的研究揭示了基底神经节环的功能。正常情况下,SNr 神经元处于紧张状态,抑制控制扫视眼球运动的丘状神经元的输出。当直接通路纹状体神经元被皮质额叶眼动区兴奋时,信噪比神经元被暂时抑制,从而使丘状神经元摆脱抑制。这允许适当的丘状神经元向眼球运动的目标发出信号,允许猴子将目光转向新的位置。运动开始于额叶视野;然而,基底神经节的强直抑制输出的功能是什么?回忆运动皮层章节在不同位置刺激猴子的运动皮层会导致刻板的动作序列,例如将手放在嘴边或采取防御姿势。许多“原始”运动程序似乎存储在皮层中,运动控制可能需要以精确的时间顺序激活这些基本运动程序,以完成复杂的运动计划。然而,在给定时间只有一个运动程序处于活动状态是很重要的,这样一种运动行为(例如,用手将食物送到嘴里)不会与冲突的运动行为(例如,用手遮住脸)竞争来自危险物体)。据认为,基底神经节通常活跃于抑制不适当的运动程序,直接通路的激活暂时解除了一个运动程序的抑制,使其能够被生物体执行。因此,基底神经节充当了能够在层次结构中执行自动程序的门。
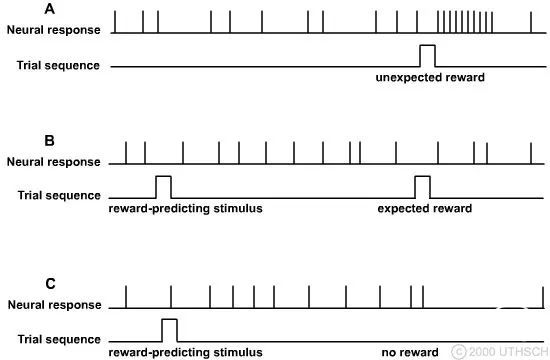
多巴胺能神经元发出意外奖励或意外缺乏奖励的信号(Hollerman 和 Schultz,1998)。(A) 如果奖励意外发生,多巴胺能神经元会迅速激发。这可能会加强导致奖励的皮层运动程序。(B) 如果猴子先前学到的奖励是由刺激预测的,则神经元放电不会改变,从而表明一切都在正常进行。(C) 如果奖励预测刺激不产生奖励,则神经元放电受到抑制。这种抑制可能会削弱没有产生预期奖励的皮层运动程序。
哪些运动程序应该在特定时刻解除抑制?基底神经节可能在了解哪些运动行为会导致对生物体的奖励方面发挥重要作用。该信息由SNc 和腹侧被盖核的多巴胺能神经元提供。猴子中这些神经元的记录表明,当猴子收到意想不到的奖励时,它们往往会做出反应,而当猴子没有收到预期的奖励时,它们往往会受到抑制。因为激活黑质纹状体通路的净效应是激发直接通路并抑制间接通路,这种多巴胺能激发模式可能参与调节直接/间接通路活动的相对平衡,以增强产生奖励结果的皮层运动程序的激发,并抑制不产生奖励的运动程序的活动。通过这种方式,可以建立倾向于奖励动物的运动习惯。
认知功能
如前所述,通过基底神经节有许多皮层环,涉及前额叶联合皮层和边缘皮层。通过这些循环,基底神经节被认为在认知功能中发挥作用,类似于它们在运动控制中的作用。也就是说,基底神经节参与选择和启用存储在这些其他皮质区域的各种认知、执行或情感程序。此外,基底神经节似乎参与某些类型的学习。例如,在啮齿类动物中,纹状体对于动物学习某些刺激反应任务是必要的(例如,如果存在刺激 A,则右转,如果存在刺激 B,则左转)。大鼠纹状体神经元的记录显示,在训练的早期,纹状体神经元在许多位置发射,而大鼠在 T 形迷宫中学习这样的任务。这表明最初纹状体参与了整个任务的执行。随着动物学习任务并变得非常擅长其表现,纹状体神经元会改变它们的活动模式,仅在试验开始和结束时才放电。看来解决这个任务的学习程序现在存储在其他地方;迷宫开始时纹状体神经元的放电可能反映了皮层中适当的运动/认知计划的启用,而迷宫结束时的放电可能涉及评估试验的奖励结果。这表明最初纹状体参与了整个任务的执行。随着动物学习任务并变得非常擅长其表现,纹状体神经元会改变它们的活动模式,仅在试验开始和结束时才放电。看来解决这个任务的学习程序现在存储在其他地方;迷宫开始时纹状体神经元的放电可能反映了皮层中适当的运动/认知计划的启用,而迷宫结束时的放电可能涉及评估试验的奖励结果。这表明最初纹状体参与了整个任务的执行。随着动物学习任务并变得非常擅长其表现,纹状体神经元会改变它们的活动模式,仅在试验开始和结束时才放电。看来解决这个任务的学习程序现在存储在其他地方;迷宫开始时纹状体神经元的放电可能反映了皮层中适当的运动/认知计划的启用,而迷宫结束时的放电可能涉及评估试验的奖励结果。看来解决这个任务的学习程序现在存储在其他地方;迷宫开始时纹状体神经元的放电可能反映了皮层中适当的运动/认知计划的启用,而迷宫结束时的放电可能涉及评估试验的奖励结果。看来解决这个任务的学习程序现在存储在其他地方;迷宫开始时纹状体神经元的放电可能反映了皮层中适当的运动/认知计划的启用,而迷宫结束时的放电可能涉及评估试验的奖励结果。

纹状体的习惯学习(Jog 等,1999)。(A) 一只老鼠被训练在 T 形迷宫中奔跑,如果听到高音就向左转以获得食物奖励,如果听到低音则向右转以获得食物奖励。(B) 在训练的早期,当老鼠开始学习任务时,纹状体神经元在迷宫的各个位置发射,尤其是在选择点。(C) 在训练后期,当大鼠掌握了任务并非常快速和准确地执行时,纹状体神经元现在仅在迷宫的开始和结束时激发。
在人类中,基底神经节似乎是某些形式的内隐记忆任务所必需的。像上面讨论的运动习惯学习一样,许多类型的认知学习需要反复试验并且通常是无意识的。一个例子是概率分类。在这种类型的任务中,人们必须学习根据属于某个类别的概率而不是任何明确的规则来对对象进行分类。在一个实验中,受试者被展示了一副带有不同符号的纸牌。每个符号都与预测下雨或阳光的一定概率相关联,受试者必须在每次试验中说出该符号是预测下雨还是阳光。因为同一个符号有时预测晴天,有时预测下雨,受试者无法设计一个简单的规则,他们一开始犯了很多错误。然而,随着时间的推移,他们开始更好地对符号进行适当的分类,尽管他们仍然经常声称是在猜测。基底神经节障碍患者在这项任务中受损,这表明基底神经节认知循环的处理在某种程度上参与了我们下意识地学习与特定刺激相关的预测结果的概率的能力。

基底神经节障碍患者在学习这种隐式概率分类任务时受到损害,他们必须根据一组 4 个提示卡预测天气(Packard 和 Knowlton,2002)。
基底神经节疾病
许多神经系统疾病是由基底神经节损伤引起的。这里将简要讨论其中两种疾病(帕金森氏病和亨廷顿氏病),以便将基底神经节相关概念与这些疾病的症状联系起来。
黑质纹状体通路与帕金森病
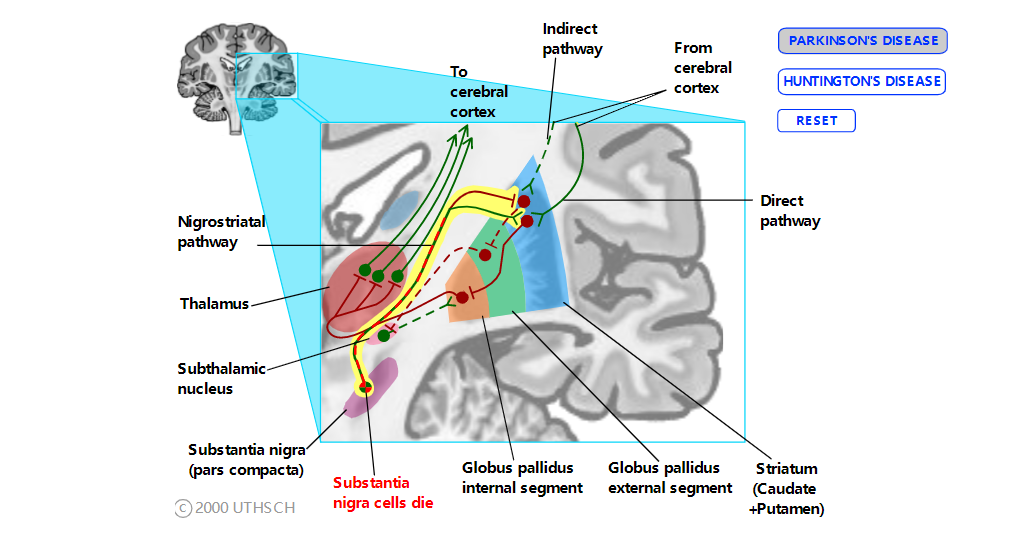
帕金森病的特征是运动缓慢或缺乏运动(运动迟缓或运动不能)、僵硬和静止性震颤(尤其是手和手指)。患者开始运动有困难,一旦开始运动就会异常缓慢。帕金森氏病的原因是丧失多巴胺能神经元在黑质致密部。根据对黑质纹状体通路对直接和间接通路影响的了解,很容易看出为什么这条通路的丧失会导致帕金森病症状的运动困难。因为黑质纹状体通路会激发直接通路并抑制间接通路,所以这种输入的丧失打破了平衡,有利于间接通路中的活动。因此,GPint 神经元异常活跃,从而抑制了丘脑神经元。没有丘脑输入,运动皮层神经元就不会那么兴奋,因此运动系统不太能够响应患者的意志来执行运动计划。
间接通路与亨廷顿病
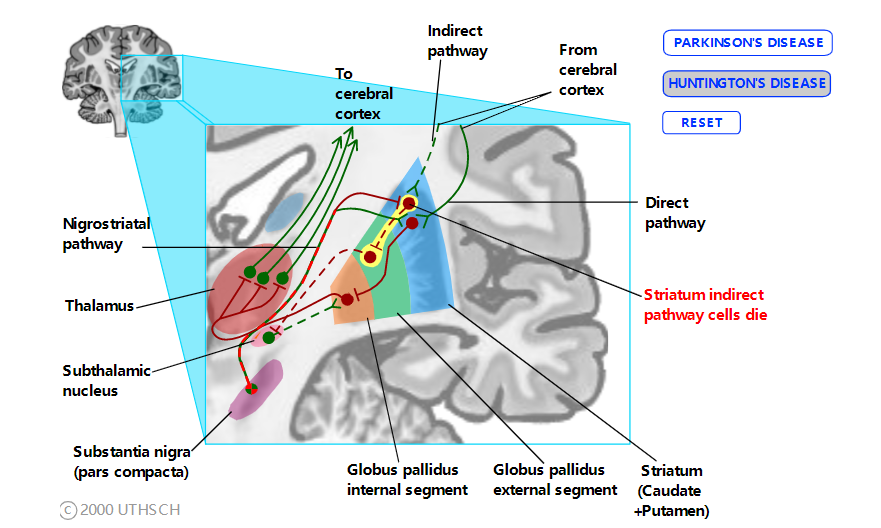
亨廷顿病的症状在许多方面与帕金森病的症状相反。亨廷顿病的特点是舞蹈动作:身体不自主的、持续的运动,尤其是四肢和面部。通常这些运动类似于适应性运动的片段,但它们是不由自主地发生的,没有行为意义。亨廷顿氏病是由间接途径中纹状体神经元的选择性丢失引起的。因此,直接途径和间接途径之间的平衡变得有利于直接途径。如果没有由间接通路提供的对丘脑的正常抑制影响,丘脑神经元会随机和不适当地激发,导致运动皮层在患者无法控制的情况下执行运动程序。
参考:
https://nba.uth.tmc.edu/neuroscience/m/s3/chapter04.html
所有分享仅供学习参考,如有错误欢迎指正。
